
生物学
独立行政法人水産総合研究センター 東北区水産研究所資源海洋部 浮魚・いか資源グループが執筆したものです。
本文中の片カッコの数字は、参考にした文献の番号です。主なものだけ引用しました。
平成26年5月1日現在の情報です。
1.サンマの分類

図1. サンマ(学名 Cololabis saira (Brevoort, 1856))
日本で食用にされるサンマは、トビウオやサヨリ、ダツなどに近い種類です。学術的(分類学的)には、ダツ目サンマ科サンマ属に属します。1) サンマ属(ぞく)の「属」とは、分類学的に最も近い種類を集めたグループのことです。さらに分類的に近い属を集めたものが「科」というグループ、さらに科を集めたのが「目」です。現在、全世界ではサンマ科の魚類は4種が知られています。2)
日本近海に分布するサンマ科の魚類は、私たちがよく目にするサンマの1種類のみです。学名(世界中で通じるラテン語の名前)では、「Cololabis saira(コロラビス サイラ)」といいます。学名のCololabisはサンマ属、sairaはサンマという種を示しており、サンマ属はサンマ1種のみです。大西洋や太平洋の南半球には「Scomberesox saurus」という「Scomberesox属」のサンマ科の1種が分布しています。サンマ科にはそのほかに2種類が知られています。
新種を見つけた時には、その種類の外見的な特徴を記録し、あらたに学名をつけて学術論文で報告されます。その学術論文がもととなって、その種は世界中に知られるようになります。サンマが新種として報告されたのは1856年です。幕末に日本に来航したペリー艦隊の学術調査隊が静岡県の下田で手に入れた標本の記録に基づいて、「ペリー艦隊日本遠征記 2巻」の中でJames Carson Brevoortという学者が報告しました。3) 学名では「Cololabis saira (Brevoort, 1856)」というように最後にこの報告者の名前を付けることもあります(省略されることも多いのですが)。当時は大西洋のサンマと同じScomberesox属の1種と判断され、学名は「Scomberesox saira Brevoort,1856」とされました。後にサンマ属(Cololabis属)に分けられたため、4) 現在のサンマの学名はCololabis saira(Brevoort, 1856)となっています。なお、報告者にカッコが付いたのは、属名が変更されたことを示しています。
「Cololabis」の語源は、ギリシア語のkolos =(短い)とラテン語のlabia = (唇)を組み合わせたものと言われています。5) また、種名の「saira」は、関西でのサンマの方言である「サイラ」から付けた名前です。サンマ属(Cololabis属)に属しているのはサンマ1種「Cololabis saira」のみです。ペリー艦隊日本遠征記によれば、この魚は「サイラウオ」と呼ばれていたと記されています。ただ、南伊豆の魚名の方言を見てもサンマを「サイラウオ」と呼んでいたという記録はなく、6) もしかすると調査隊にこの名前を教えた日本人は、関西の出身者だったのかもしれません。
2.サンマの分布
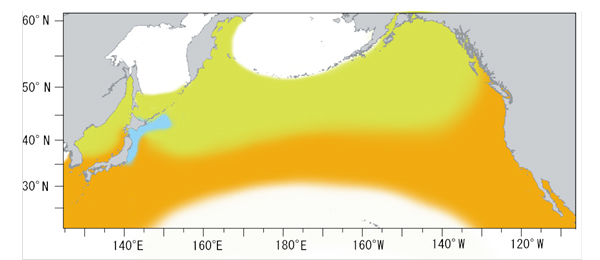
図2.サンマの分布域。黄緑とオレンジ色および水色がサンマの分布する海域。オレンジは産卵
場、水色は日本漁船の漁場を示す。
サンマは北太平洋の広い範囲に分布しています。分布域は、東西方向には北太平洋の日本海を含む日本沿岸から南米のカリフォルニア沖まで、南北方向分布には、北緯20゚から北緯60゚近くまでの亜熱帯域から亜寒帯域まで広がっています。2) しかし、1年を通してこの海域全域に分布しているわけではなく、夏は分布域の北側に、冬は南側に生息しています。このような南北の回遊をするのは、夏には北の海域に行って豊富なプランクトンを食べ、秋から冬にかけて南の海域にある産卵場に向かうためです。7) またサンマは単に南北に回遊するだけではなく、東西方向にも広く移動することが知られています。8) (回遊の項参照)
サンマは海の表面近く(表層)に分布し、あまり深くもぐることはありません。道東や千島沖の調査では、昼間に水深10~15m付近を遊泳していることが確かめられましたが、9) 房総沖では水深70m位で魚群探知機で確認されたという報告もあります。10) 海の水温は表層で高く、深くなると冷たくなります。表面の水温が高く暖かい層の厚さは場所によって違い、北で浅く南で深くなっています。サンマは表面の暖かい層に生息しているため、生息の最深深度は南北で異なるようです。
このように海の表面近くに生息する魚類を、浮魚(うきうお)と呼びます。マイワシやマサバ、カツオやまぐろ類なども浮魚類です。浮魚類の体色は背中が濃い青、腹側が銀色になっていることが多いのですが、この体色は保護色と考えられています。上から鳥などの捕食者に狙われたときには海の色に溶け込み、下から大型の魚類などに狙われた時には降り注ぐ光に紛れ込んで見つかりにくくするためといわれています。11)
3.成熟と産卵
サンマはほぼ1年中産卵しています。しかし、産卵量は季節的に変化し、太平洋側では冬に最も盛んに産卵が行われており、夏はわずかです。12) 水産総合研究センター中央水産研究所では、毎年1~3月に本州南側の太平洋の黒潮が流れる海域でサンマの子供(仔稚魚;しちぎょ)の量を調査しています。この調査では、沿岸よりも黒潮の流れの本流よりも沖の海域でより多くのサンマが採集されています。13) さらにその東側でもサンマの仔稚魚は採集されており、12) 北アメリカカリフォルニア沖でも仔稚魚が採集されている14) ことから、サンマの産卵場は分布域の南側全体に広がっていると考えられています。
サンマの卵はやや楕円(だえん)形で、卵径は1.32~2.08mmくらいです。卵には纏絡糸(てんらくし)と呼ばれる細い糸がついており、流れ藻などの漂流物に絡みついています。近縁種のサヨリやトビウオの一部の種類はサンマによく似た卵を産みますが、卵の大きさや形、纏絡糸の付き方で区別できます。10,15) しかし、仔稚魚が多数採集される冬の太平洋では流れ藻がほとんどなく、どこに卵を産み付けているのかはよくわかっていません。
卵は15℃で14日、20℃では10日ほどでふ化します。16) 孵化時にはすでに一人前のサンマと同じく背中側は青、腹には白または銀色の色素が沈着しており、卵黄は少し残っている程度です。17) 多くの魚種では孵化時にはほとんど透明で体の色素はごくわずかで大きな卵黄をつけていますが、サンマでは体色も成魚とほとんど同じで、卵黄もかなり吸収されるなど、卵の中で仔魚の発育が他の魚種よりも進んでから孵化します。
産卵を開始する体長は、一般的には体長25cm以上18) ですが、まれに20cm位で産卵している19,20) こともあります。産卵期には数日おきに産卵を繰り返します。そのため、産卵中のサンマの卵巣中にはいろいろな大きさの発達段階の異なる卵が見られます。18) 飼育実験では産卵が3カ月以上続きましたが、21) これは餌を十分与えた時の結果なので、野外における産卵の継続期間はもう少し短いと考えられます。野外で採集されたサンマの卵巣中の卵を数えた結果から、1回の産卵で500~4000個ぐらい卵を産むと見積もられています。21,22) 1回の産卵期には、30回以上産卵を繰り返すと推定されています。23)
しかし、野外で採集したサンマを見てみると、25cm以上のサンマがすべて産卵しているわけではありません。また、産卵を続ける期間も、産卵を始める季節やサンマの年齢、栄養状態によって変化すると思われ、実際にサンマが一生の間にどれくらい卵を産むかはよくわかっていません。さらに、生み出された卵もどこにあるのかなど、野外における産卵の実態については、よくわかっていないことが数多くあります。
4.サンマの餌、天敵
サンマは生まれた直後から肉食性で、カイアシ類やオキアミ類などの小型の甲殻類(こうかくるい;エビやカニ、ミジンコなどを含むグループ)を食べています。24-27) サンマの生態を観察・研究するために、水産総合研究センター北海道区水産研究所では実験用にサンマの飼育を行ったことがありますが、28,29) この時には先に水族館でサンマの飼育と展示に成功していた「ふくしま海洋科学館(アクアマリン福島)」30) の方法を参考に、仔稚魚にはワムシやアルテミア、冷凍したカイアシ類、成魚には配合餌料、魚やエビ類をすりつぶしたものを、餌として与えていました。
サンマは胃がないので食べたものを蓄える大きく膨らんだ袋状や折れ曲がった構造はなく、口から肛門まで消化管が一直線になっています。19,31) 飼育下では餌の量は最大で体重の13%ほどに達します。19) また、消化のスピードは速く、3時間くらいで2/3以上の餌が排出されることがわかっています。19) 野外では日没直後に消化管中の餌が多いことからこの時間帯に餌を摂るようです。32)
一方、サンマを食べる生物として、ミンククジラなどの鯨類、33) ハイイロミズナギドリ、34-36) ウトウ37) などの鳥類、ギンザケ、38) ビンナガ39) などの大型魚類が知られており、沖合に生息する高次捕食者(食物連鎖で上にいる動物)の重要な餌となっています。
5.年齢と成長
サンマの寿命は約2年です。40) 他の小型浮魚類の寿命が、マイワシで最大8年、マサバで11年である41) のと比べると、非常に短いのが特徴です。漁期中に漁獲されるサンマではおおむね29cmより小さいものが1歳未満、それ以上は1歳魚です。40) 普段私たちが食用とするサンマは体長が30cm前後ですが、これらは多くの場合1歳魚です。最大体長は35cmくらいですが、ごく稀に40cm近くになるものが出現することがあります。42) 成長のはやさは生まれた時期43,44) や、育った海域45) によっても変化することが知られています。
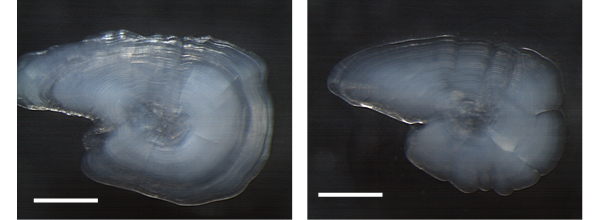
図3. サンマ耳石 白い線は 0.5mm を示す。
現在サンマの年齢は、2つの方法によって調べられています。いずれも、頭の中にある耳石(じせき)をいう炭酸カルシュームの塊を調べる方法です。サンマの耳石は体長30cm位でもその長さが2mmほどと小さく厚さも薄いので、初めて探す人ではなかなか見つけられません。耳石の中心にある核から同心円状に成長し、体長が大きくなるとともに耳石も大きくなります。
年齢を調べる第1の方法は、この耳石にできた年輪を数えることです。年輪は1年に1回、秋にでき始め、46) 2月ごろには形成が終わります。47) 普通の光学顕微鏡(学校の実験で使うような顕微鏡)で上から光を当てて耳石を観察すると、1歳魚では黒い帯状の年輪が見えます。この方法は耳石を丸ごと観察するだけで簡単にみえるので、たくさんのサンマの年齢を調べることができます。しかし、わかるのは年齢だけで、ふ化した季節まではわかりません。また、年輪の半径は最初に秋を迎えた時の体長の目安になります。
第2の方法は、耳石中に毎日形成される細かい輪紋(耳石日周輪)を観察する方法です。43,44,48) 日周輪は多くの魚類で日齢(生まれてからの日数)を調べるのに使われています。

図4. サンマの耳石の電子顕微鏡写真
小さいサンマでは光学顕微鏡でも数えることができますが、大きくなると耳石が厚くなり綸紋の間隔も狭くなるので、耳石を半分に削ったり、観察に電子顕微鏡を使う必要があるなど、観察には非常に手間がかかります。そのため、たくさんの個体について耳石日周輪から日齢を調べるのは難しいのですが、孵化時期や毎日の成長の変化が綸紋の間隔から推定できる43,44) など、過去の成長の履歴を探る上で欠かせない手法となっています。この耳石日周輪は、秋に年輪ができ始めるとその先は数えることができません48) が、その成長パターンから、おおよその孵化時期も推定することができます。44,45)
6.資源量と資源調査
北太平洋に広く分布しているサンマですが、どれくらいの量のサンマがいるのでしょうか?水産総合研究センター東北区水産研究所では2003年から毎年6~7月に、日本の近海から西経165°までの海域で、調査船によってサンマがどれくらいいるか(資源量)を調べる調査を行っています。
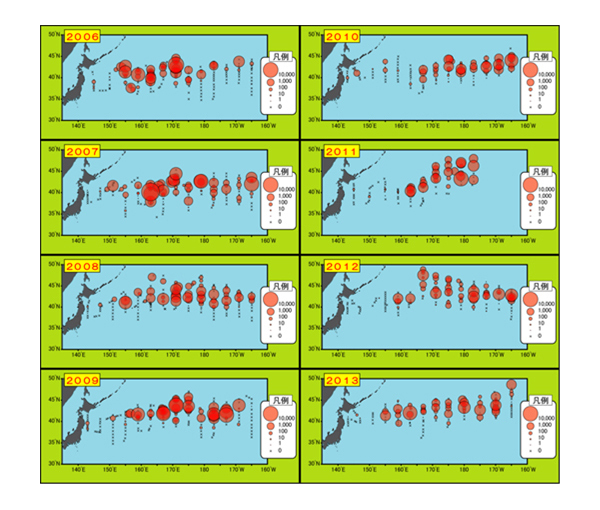
図5.2006年から2013年まで毎年6~7月に行われたサンマ漁期前調査の結果。丸の大きさは採
集されたサンマの数を示す。2010年以降、日本近海から東経160度(160°E)までの領域で
サンマが少なくなっているのがわかる。
西経165°は日付変更線を越えたハワイの方が近い海域です。この調査でサンマの分布や回遊、年ごとの資源量の変化などがわかってきました。49) 調査海域全体では、サンマの量は2003年にもっとも多く502万トン、2012年に最も少なく198万トンと推定され、その差は2.5倍以上ありました。
サンマの寿命は2年と短く、スーパーなどで一般の消費者向けに売られているものは1歳魚がほとんどです。このことは、サンマの資源は2年でサンマ資源は完全に世代が交代し、毎年どれくらいの数のサンマが育つかによって、その資源量が大きく変化することを示しています。つまり、毎年どれくらいの卵が生み出され(これは親の数と、その親がどれくらいの卵を産卵できるかによります)、そのうち生き残るサンマの割合(これは海の状態(海洋環境)が大きく関係すると考えられています)によって、資源量が増えたり減ったりします。人間では少子化の影響が数十年たって、少子化した世代が成人し働き盛りとなってから深刻な問題となりますが、寿命が短いサンマでは少子化が翌年には深刻な問題となるのです。
サンマをはじめ多くの魚類はたくさんの卵を産みますが、そのほとんどは大人になる前に死んでしまい、ごく少数のみが生き残って次の世代となります。1匹の雌から数万個生み出されたサンマの卵のうち、2匹のサンマが残れば次の世代の数は同じになります。ところが数万のうちの4匹が残れば次世代のサンマの数は2倍に、逆に1匹しか残らなければ半分になってしまいます。また、親の数が前の年より半分になれば生み出される卵は半分になり、前の世代と同じ割合のサンマが生き残っても次世代の数は半分になってしまいます。このように多くの子供を産んでごくわずかの子供が生き残る生物では、生き残りの率や親の数が少し変わっただけで、次世代の数は大きく変化します。また、寿命が短いほどその変化が急に現れます。ですから、今年たくさんサンマが獲れたとしても、2年後にはどうなるかはまったくわからないのです。このような種類では将来の資源量を予測することが難しく、サンマ資源の量を正確に把握するためには、毎年の資源量調査が欠かせません。また、調査ではサンマの数が変化する原因も明らかにするため、海の水温や餌となる生き物の量も調べています。
また、この調査では漁期が始まる前のサンマが、どこにいるかもわかりました。日本の漁船は8月以降日本の近海でサンマを漁獲していますが、漁期前調査を行っている6~7月には日本のごく近海にはサンマが少なく、沖合の東経150°より東に多く分布していました。49) 調査時期における東経150度より沿岸側の資源量は漁期中に漁獲されるサンマの水揚げ量よりもずっと少なく、漁期に漁獲されるサンマは漁場よりも沖からやってくると考えられました。50) ではどのくらい沖からやってくるのでしょうか?
6~7月の漁期前調査時に海域ごとに1歳魚の体長を見ると、東経160~170度を境に日本に近い海域では体長は大きく、沖合で小さくなっています。耳石年輪の半径を見ても同様に沿岸の方が大きくなっています。ところが、孵化時期は沿岸側でも沖合でも顕著な差はなく、1年目の成長が海域によって違うようです。45) 一方、10月以降に日本近海で漁船が漁獲したサンマの耳石の年輪を計測すると、6~7月に東経160度より沖にいたサンマと同じくらいの大きさです。つまり、漁期中に漁場にやってくるサンマは東経160度より沖合から来遊するものがメインになっていると考えられます。8)
6~7月の調査結果から、サンマの資源量を3つの海域(調査海域を東経162度、西経177度で3つに区分)に分けて調べています。資源量は、東経162~西経177度までの安定しているものの、2010年以降東経162度より沿岸側の日本にもっとも近い海域の資源量が著しく減少しています(上図を参照)。また、沿岸側の資源量減少に伴い、2010年以降漁期解禁直後の8月から9月上旬の漁場が遠くなり魚群も薄くなっていることから、解禁直後の水揚げ量が減少しています。9月中旬以降は漁獲量が伸びていますが沖合から来遊してくるためと考えられます。49)
7.サンマを漁獲する国々
FAO(国際連合食糧農業機関)の統計51) によると、2010年現在、サンマを漁獲している国は日本、ロシア、台湾、韓国です。長らく漁獲量のほとんどは日本によるものでしたが、2000年以降台湾の漁獲量が急激に伸び、2013年には日本の漁獲量を超えたようです。このほか中国も近年公海でのサンマ漁業に参入しました。漁場や漁業を行う期間は国によって異なり、日本とロシアが8月から12月まで主に近海の排他的経済水域(EEZ;陸から200海里(約370km)までの海域)内で漁獲しているのに対し、台湾と韓国は主にその外側(公海)で6月から12月まで漁業を行っています。
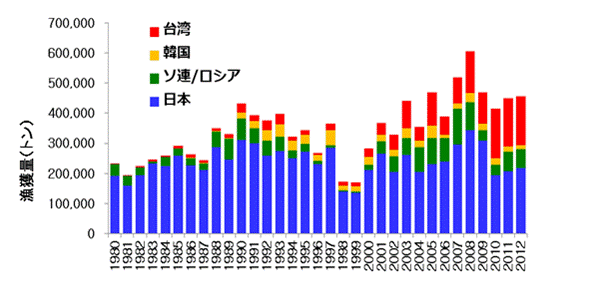
図6.1980年から2012年まで各国の年別のサンマの漁獲量の推移。日本とロシアは近海(EEZ内)
での漁獲が主、台湾と韓国は公海での漁獲が主体となっている。1990年までは日本とロシア
の漁獲量が90%以上を占めたが、2000年代以降は台湾の漁獲量が増加している。FAO統計の
ほか、各国の研究者への聞き取り結果を加えて作成した。
日本の漁船の大きさは200トン未満で、漁獲したサンマは氷蔵して港に運んでいます。一方、台湾の漁船は1000トンほどで漁期を通して漁場にとどまり、漁獲したサンマは船上で箱詰めされて冷凍保管されたあと、運搬船で港に運ばれます。52) このため、日本の漁船は遠くまで行くことが難しく、2010年以降サンマの分布が沖に移動し近海に来遊する魚群が減少したため、その影響が漁獲量に直接影響しています。一方、台湾の漁船は魚群が沖に移動しても漁船も移動するためその影響は少なく、近年逆に漁獲量が増加しています。ただし、サンマ全体の資源量は減少しているため、漁獲率(資源全体に対する漁獲量の割合)は年々増加し、過去は10%以下だったものが2012年は20%を越えています。
しかし、先に述べたように沖にいるサンマもやがて日本近海に来遊するので、漁場は違ってもサンマの漁業国は同じ群(資源)を利用しています。そのため、サンマの漁業国は同じ資源を利用しているという意識を明確にし、将来にわたって安定してサンマ資源を利用(続的利用)できるように、漁獲量や漁獲方法を考える必要があります。かつて日本がサンマの漁獲量の大部分を占めていた時代には、公海域では分布していたサンマは漁獲されず、結果的に禁漁区になっていました。ところが、近年のように公海においても多くのサンマが漁獲されるようになると、資源の動向にどのような影響が出るかを過去の記録だけから推測するのは難しいのです。
このような状況の中、「北太平洋における公海の漁業資源の保存及び管理に関する条約(略称:北太平洋漁業資源保存条約)」という国際条約が間もなく発効され、53) 近いうちにこの枠組みの中で国際的にサンマの資源管理が行われるようになります。ペリー艦隊の調査隊の記録をもとにサンマが世界に紹介されてから150余年、サンマ資源は今後、国際的にも大きな注目を浴びてゆくことになります。
8.参考文献
1.サンマの分類
1)中坊徹次 編 (2013)「日本産魚類検索 全種の同定 第3版」.東海大学出版会,秦野,2428pp.
2)Hubbs, C. L. (1980) Revision of the sauries (Pisces, Scomberesocidae) with descriptions of two new genera and one new species. Fish. Bull. US, 77, 521-566.
3)Brevoort, J. C. (1856) Notes on Some Figures of Japanese Fish: Taken from Recent Specimens by the Artists of the US Japan Expedition. In Narrative of Commander M. C. Perry's Expedition to Japan 2:253-288. (原著は下記のURLで図版とともに見ることができます。https://archive.org/details/notesonsomefigur00brev サンマの解説は29ページ、図版は53ページ)
4)Gill, T. (1895) The families of synentognathous fishes and their nomenclature. Proceedings of the United States National Museum. 18:167-178.
5)Fishbase http://www.fishbase.org/summary/303
6)松岡玳良 (1975) 伊豆半島各地の魚の方言(I).伊豆分場だより,第180号,19-25.
2.サンマの分布
7)福島信一 (1979) 北西太平洋系サンマの回遊機構の綜観的解析.東北区水産研究所研究報告,41,1-70.
8)Suyama, S., Nakagami, M., Naya, M.,Ueno, Y. (2012) Migration route of Pacific saury Cololabis saira inferred from the otolith hyaline zone. Fisheries Science, 78(6), 1179-1186.
9)和田時夫,北片正章 (1982) サンマ中層流し網における採集試験結果と昼間の行動について.北海道区水産研究所報告,47,11-22.
10) 堀田秀之 (1964)「サンマ資源」.日本水産資源保護協会,東京,96pp.
11) 木村 重 (1978)「魚の生態」.刊々堂,320pp
3.成熟と産卵
12)Watanabe, Y., Lo, N. C. (1989) Larval production and mortality of Pacific saury, Cololabis saira, in the northwestern Pacific Ocean. Fishery Bulletin, 87(3), 601-613.
13) Takasuka, A., Kuroda, H., Okunishi, T., Shimizu, Y., Hirota, Y., Kubota, H., Sakaji, H., Kimura, R., Ito, S., Oozeki, Y. (in press) Occurrence and density of Pacific saury Cololabis saira larvae and juveniles in relation to environmental factors during the winter spawning season in the Kuroshio Current system. Fisheries Oceanography.
14)Ahlstrom, E. H., & Stevens, E. (1976) Report of neuston (surface) collections made on an extended CalCOFI cruise during May 1972. Calif. Coop. Oceanic Fish. Invest. Rep, 18, 167-180.
15) 遊佐多津雄 (1958) サンマとサヨリの魚卵と稚魚の主な相違点について.北水試月報,15,249-256.
16) 堀田秀之,福島信一 (1963) 東北海区に於けるサンマ資源の数量変動に関する研究.東北区水産研究所研究報告,23,73-84.
17) Yusa, T.(1960) Embryonic development of the saury Cololabis saira (Brevoort)東北区水産研究所研究報告,17,1-14.
18) Hatanaka, M. (1956) Biological studies on the population of the saury, Cololabis saira (Brevoort) Part 1 Reproduction and growth. Tohoku Journal of Agricultural Research. 6, 227-312.
19) 堀田秀之 (1958) 飼育実験によるサンマの成長について.東北区水産研究所研究報告,11,47-64.
20) 菅間 慧一 (1957) 耳石の性状からみたサンマのポピュレーション構造-I.北海道区水産研究所報告,16,1-12.
21) 小坂淳 (2000) 北西太平洋におけるサンマの生活史とそれに基づく資源変動の考察. 東北区水産研究所報告, 63, 1-96.
22) 巣山哲 (2002) 北太平洋におけるサンマCololabis saira (Brevoort)の年齢,成長および成熟に関する研究.水産総合研究センター研究報告,5,68-113.
23) 栗田豊 (2002) 環境変動に対するサンマの繁殖特性の応答.我が国周辺海域における漁業資源の変動予測技術の開発.「農林水産技術会議事務局編」,農林水産省農林水産技術会議事務局,東京,60-63.
4.サンマの餌、天敵
24) 堀田秀之,小達和子 (1956) サンマの食餌構成と摂餌行動に就いて.東北区水産研究所研究報告, 7, 60-69.
25) 小達和子 (1977) サンマの食性について. 東北区水産研究所研究報告, 38, 75-101.
26) 高 幸子,北片正章,和田時夫 (1980) 千島列島南東水域における7 月のサンマ餌料生物と動物プランクトンの鉛直分布について.北海道区水産研究所報告,45,15-41.
27) Sugisaki, H.,Kurita, Y. (2004) Daily rhythm and seasonal variation of feeding habit of Pacific saury (Cololabis saira) in relation to their migration and oceanographic conditions off Japan. Fisheries Oceanography, 13(s1), 63-73.
28) Nakaya, M., Morioka, T., Fukunaga, K., Murakami, N., Ichikawa, T., Sekiya, S., Suyama S., Shimizu, A. (2011) Verification of growth dependent survival in early life history of Pacific saury Cololabis saira using laboratory experiment. Environmental biology of fishes, 92, 113-123.
29) 巣山 哲 (2013) 飼育下におけるサンマの産卵生態. 「漁業資源の繁殖特性研究-飼育実験とバイオロギングの活用 (水産学シリーズ175) 」.栗田 豊,河邊 玲,松山倫也編,恒星者項性格厚生閣,東京,92-105.
30) 津崎順 (2000) サンマの飼育と展示- Ⅰ. AMF NEWS, 2(2), 2-3.
31) 長澤和也 (1987) サンマ.「魚類解剖図鑑」落合明編,緑書房,東京,123-128.
32) 栗田豊,杉崎宏哉 (2004) サンマ日間摂餌量の季節および体長による変化. 水産海洋研究, 68, 133-141.
33)Tamura, T.,Fujise, Y. (2002) Geographical and seasonal changes of the prey species of minke whale in the Northwestern Pacific. ICES Journal of Marine Science, 59, 516-528.
34) Ogi, H. (1984) Feeding ecology of the Sooty Shearwater in the western subarctic North Pacific Ocean. Marine Birds: Their Feeding Ecology and Commercial Fisheries Relationships, ed.by D. N. Nettleship et al. Canadian Wildlife Service Special Publication, Ottawa, 78-84.
35)Gould, P., Ostrom, P.,Walker, W. (2000) Foods, trophic relationships, and migration of Sooty and Short-tailed Shearwaters associated with squid and large-mesh driftnet fisheries in the North Pacific Ocean. Waterbirds, 23,165-186.
36) Shiomi, K., Ogi, H. (1992) Feeding ecology and body size dependence on diet of the sooty shearwater, Puffinus griseus, in the North Pacific. In Proceeding of National Institute of Polar Research Symposium, Polar Biology 5, 105-113.
37)Vermeer, K. (1980) The importance of timing and type of prey to reproductive success of Rhinoceros Auklets Cerorhinca monocerata. Ibis, 122, 343-350.
38) 佐藤忠勝,平川英人 (1976) 北西太平洋におけるギンザケの食性に関する研究.福島県水産試験場研究報告,4,25-31.
39) 二平 章 (1988) 北太平洋・天皇海山周辺海域に分布するビンナガTunnus alalunga (Bonna Terre)とサンマCololabis sairaの補食者・被食者関係.茨城県水産試験場研究報告26.125-136.
5.年齢と成長
40) Suyama, S., Kurita, Y., Ueno, Y. (2006) Age structure of Pacific saury Cololabis saira based on observations of the hyaline zones in the otolith and length frequency distributions. Fisheries Science, 72(4), 742-749.
41) 平成25年度 水産庁増殖推進部・独立行政法人水産総合研究センター (2013) 「わが国周辺水域の漁業資源評価 第1分冊」.水産庁,東京,
42) 今井義弘 (1986) 1985年秋に漁獲された超特大サンマについて第35回サンマ研究会報告.248-251.
43) 根本豊,栗田豊,大関芳沖。本間隆之,林崎健一,井田齋 (2001) サンマ耳石微細輪紋の形成様式. 東北区水産研究所研究報告, 64, 69-78.
44) Kurita, Y., Nemoto, Y., Oozeki, Y., HAYASHIZAKI, K. I., Ida, H. (2004) Variations in patterns of daily changes in otolith increment widths of 0+ Pacific saury, Cololabis saira, off Japan by hatch date in relation to the northward feeding migration during spring and summer. Fisheries oceanography, 13(s1), 54-62.
45) Suyama, S., Nakagami, M., Naya, M., & Ueno, Y. (2012) Comparison of the growth of age-1 Pacific saury Cololabis saira in the Western and the Central North Pacific. Fisheries science, 78, 277-285.
46) Suyama, S., Oshima, K., Nakagami, M.,Kawabata, A. (2011) Seasonal changes in otolith and somatic growth in age-0 Pacific saury Cololabis saira. Fisheries Science, 77, 223-233.
47) 堀田秀之 (1960) 鱗・耳石によるサンマのポピュレーション構造の分析とその成長.東北区水産研究所研究報告,16, 41-64.
48) 巣山哲, 桜井泰憲, 目黒敏美, 島崎健二 (1992) 中部北太平洋におけるサンマ Cololabis saira の耳石日周輪に基づく年齢と成長の推定. 日本水産学会誌, 58, 1607-1614
6.資源量と資源調査
49) 中神正康 (2013)「近年のサンマ資源と漁業の動向」.東京水産振興会,東京,60pp.
50) 伊藤進一,大野創介,岸道郎,尹錫鎭,巣山哲,中神正康,筧茂穂,亀田卓彦,安倍大介 (2014) サンマ産卵回遊時の能動的遊泳を考慮したサンマ成長・回遊モデルの高度化第62回サンマ等小型浮魚資源研究会議報告303-309
7.サンマを漁獲する国々
51) FAO HP. http://www.fao.org/fishery/statistics/global-capture-production/en
52) 上野康弘 (2010) 待ったなしのサンマ漁の国際的資源管理 AFCフォーラム9月号,7-10.
53) 外務省HP. http://www.mofa.go.jp/mofaj/gaiko/page24_000002.html



